生物学能基本原理与能量代谢
生物学能基本原理
自由能与平衡常数
-
基本概念与原理
- 能量//生物能(环境–>生物–>环境)//热力学能
- 热力学第一定律(能量守恒)//热力学第二定律(熵增原理)
-
△G = △H - T △S = - 2.303RT*lg(K_eq)
- △G0为负值,K_eq > 1,反应趋向完全,是放能反应
- △G0为正值,K_eq < 1,反应是不利的,是吸能反应,标准情况下,几乎没有产物生成
-
在给定的环境条件下,ΔG的影响因素
- 电子供体与电子受体之间的标准还原电势差 (ΔEm,7)
- 电子供体与电子受体的绝对浓度
- 电子供体与电子受体的氧化/还原水平
-
ATP是生物能的通量
- 2类合成反应(ADP+Pi//2ADP–>ATP+AMP,后者k_cat更大)
- 能荷的定义(总是维持在0.8-0.85,此时ATP的水解和合成活性居中)
- 为什么是ATP?unexplained biological dichotomies//随机稳定
ATP与生命起源
- 原始生命起源于碱性喷口vent Lost City Hydrothermal Field
-
这里的化学反应和能量变化与细胞内的环境类似
-
生命体系是低熵的,因而需要更大的负焓(原始汤中的原始环境往往高熵低焓,而碱性喷口的环境不是)
-
与火山口相比,碱性喷口的活性周期长、温度低、pH~8,更符合生理环境
-
该环境下由CO+H2+NH4+—>氨基酸、糖的过程是放热的,因而有助于生物体系减少自身碳源的合成(矿物催化剂)
-
环境中发生着CO2的还原固定反应给了细胞形成基因、蛋白机器的可能——电子歧化反应与离子梯度
- #QUESTION:H2/H+的能量变化?歧化的电子流向以及所带来的能量变化!结合反应的物质变化在想想
- 动力学上为什么可以用H2还原CO2
- 标准状况下,CO2/HCOOH E0=-430mV、2H+/H2 E0=-414mV、若以H2作为电子供体(正极),则H2+CO2—>HCOO-+H+ ΔE=(E+)-(E-)=-16mV<0,根据能斯特方程反应不自发;因而1988年就有人认为生命体不可能用H2还原CO2
- 但是,根据能斯特方程,在碱性条件下,H电对的电极电势下降比C快,因而使得这个还原反应在动力学上可以发生!
- 在碱性喷口处由于区室化的形成,从而形成了局部的碱性环境和质子梯度,从而给H2还原CO2提供了可能
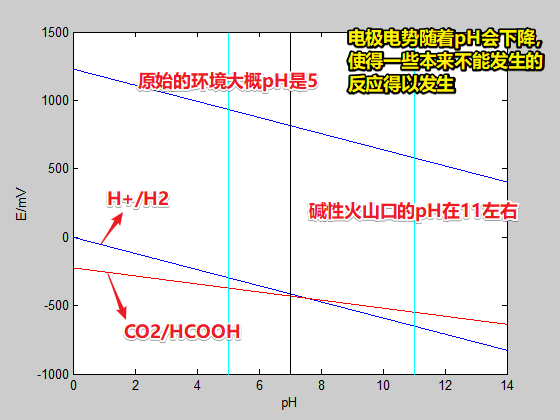
- 环境中发生的CO2还原给细胞创造了有机物环境,使得细胞可以有着先天利用有机物(如氨基酸)的优势,从而逐渐演化出了利用氨基酸形成蛋白,形成基因装置的可能
-
基因、蛋白等有机物的形成给生物膜形成奠定了基础,也使得生物膜的透性越来越低,从而形成了质子梯度
- 起初,在细胞膜透性较好时,细胞可以利用热泉的碱性水环境来平衡内部H+,维持碱性环境
- 而内外环境的质子梯度建立,可以被利用产生能量,即ATP合酶
- 但随着膜的逐渐完善和封闭,细胞必须解决H+的平衡问题以提供自身一个能量来源(H2还原CO2放能产生ATP)
- 原始状态的H pump的能量利用效率可能不高,因而必然会有大量能量用于维持质子梯度,而用于生物量积累的能量则减少
- #QUESTION:细菌如何解决这个质子梯度的问题?
-
能量的产生方式
底物磷酸化
- 从能源物质分解代谢产生的高能化合物转移磷酸基团到ADP,形成ATP的过程
- 效率较低;涉及到的酶在不同生物中分布不广泛
- 有证据表明,底物磷酸化相对化学渗透的能量产生方式,可能是更进化的
- 虽然一般认为结构更精巧、功能更高效即更进化,但也并不绝对
- 化学渗透所利用的ATP合酶普遍存在于各类生物(包括LUCA)中
- 底物水平的磷酸化却并不在生物中十分常见,其所涉及的酶没有显示出很原始的迹象
化学渗透
伴随着一系列氧化还原反应,电子沿电子传递链传递,质子泵出膜外,造成膜内外的质子梯度(化学势能)和电荷梯度(电势能),形成质子动力
基于视紫红质的化学渗透【不依赖电子传递链!】
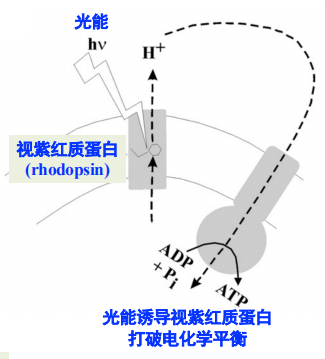
- 1973年首先在嗜盐古菌中发现盐沼盐杆菌 (Halobacterium halobium)
- 低氧条件下,光照会诱导表达更多的视紫红质蛋白,发色团是视黄醛 (retinal;维生素A醛),属于类胡萝卜素
- 视黄醛共价地结合在色素蛋白上,而色素蛋白嵌入细胞质膜中,从而使视黄醛在膜的中部
- 在视黄醛存在的情况下,光照引起含有视紫红质蛋白的大肠杆菌悬浮液 pH 降低
- 在海洋浮游细菌独岛菌属Dokdonia sp. strain MED134中发现,光照可以促进视紫红质蛋白编码基因的转录,使细胞形态发生变化,并促进其生长
- 编码基因可以在原核生物间水平转移,但局限在(海洋)透光区的微生物。
氧化还原与电子传递原理
-
氧化磷酸化: ATP的形成是基于电子传递相偶联的磷酸化
- 膜内外出现质子梯度(化学势能)和电荷梯度(电势能),形成质子动力 (健康生长的细胞约150 mV)
- 质子动力推动质子沿质子和电荷梯度穿过膜进入膜内,这种质子流是放能的,在膜上ATP合酶的作用下,使ADP磷酸化成ATP
-
氧化还原反应:电子从电子供体 (donor) 转移到电子受体 (acceptor) 的反应
- 氧化还原电对(couple):一对电子供体和受体
- 反应的平衡常数E_0:衡量供体失电子的能力或受体得电子能力; E_0’是pH=7.0的还原电势
- 电对还原电势越低,其还原物是越好的电子供体;电对还原电势越高,其氧化产物是越好的电子受体
- 好的电子供体/受体反应,其反应越易自发,其自由能越负,产能越多
-
电子载体
-
NAD+ 烟酰胺腺嘌呤二核苷酸
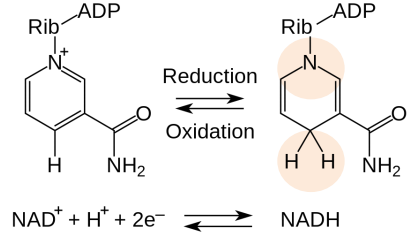
- 烟酰胺腺嘌呤二核苷酸的烟酰胺环从供体获得两个电子和一个质子
- 烟酰胺腺嘌呤二核苷酸磷酸(NADP+)类似
-
FAD 黄素腺嘌呤二核苷酸
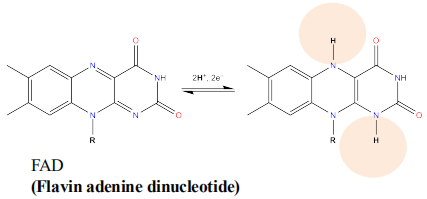
- 黄素腺嘌呤二核苷酸(FAD)的异咯嗪环从供体获得两个电子和两个质子;
- 黄素单核苷酸(FMN)类似;含有FAD和FMN的蛋白质叫黄素蛋白
-
CoQ 泛醌
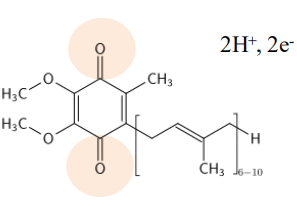
- 泛醌可从供体获得两个电子和两个质子
-
血红素
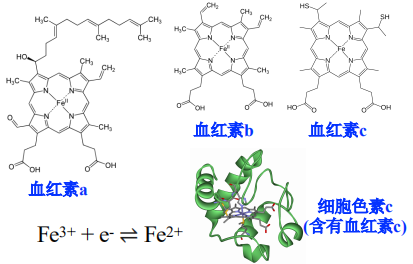
- 细胞色素(cytochrome)中,铁原子参与血红素或其他类似卟啉环的组成,由蛋白质和铁卟啉环组成
- 通过Fe原子的可逆的氧化还原反应,每次传递一个电子
-
Fd 非血红素铁蛋白(含铁原子但无血红素的电子传递蛋白)
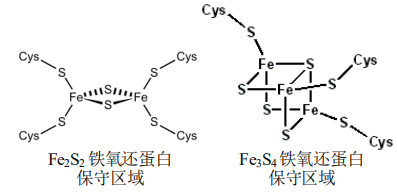
- 铁氧还蛋白(ferredoxin),虽然不含血红素,但仍有可逆的氧化还原反应。
- Fd可分为Fe2S2、Fe4S4、Fe3S4等类型
-
-
电子载体的选择:能产生更多能量的电子受体被优先利用!
-
电子传递系统
-
基本特点
- 电子从电子供体向最终电子受体的传递,要通过一系列的载体,这些电子载体组成的系统
- 第一个载体有最负的E0’,每一个接下来的载体的E0’都依次升高
- 存储在发动电子流的氧化还原电对里的势能释放,用来形成ATP
-
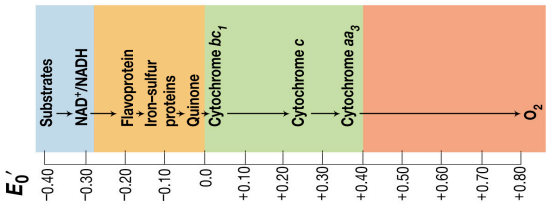
亚细胞定位(线粒体和叶绿体都起源于革兰氏阴性菌)
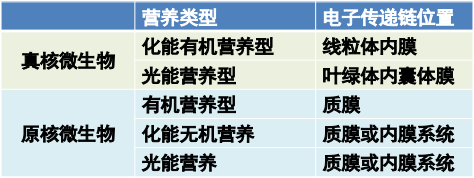
- 真核生物中化能有机营养型微生物的电子传递链在线粒体内膜
- 从NAD+/NADH,黄素单核苷酸FMN,辅酶Q,细胞色素bc1, 细胞色素c, 细胞色素aa3,还原电势越来越高
-
氧化还原底物的多样性影响能量产生的多少
- 电子供体与电子受体之间的标准还原电势差 (ΔEm,7)
- 电子供体与电子受体的绝对浓度
- 电子供体与电子受体的氧化/还原水平
-
-
【电子传递的模型】E·coli有氧呼吸电子传递链
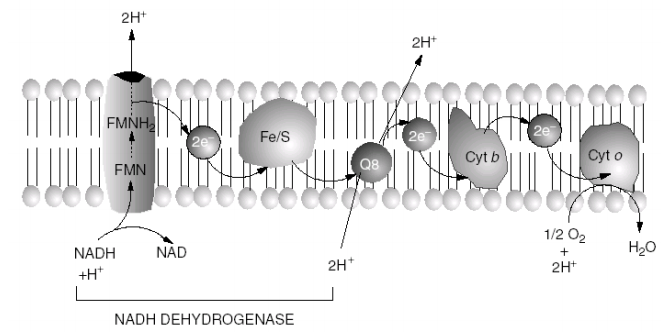
-
呼吸链:NADH—>FMNH2—>Fe/S—>Q_B—>Cyt b—>Cyt o—>O2 (第一个载体有最负的E0’,每一个接下来的载体的E0’都依次升高)
-
两种末端氧化酶
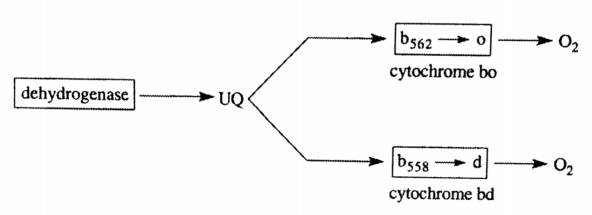
-
bo-type (对数期、高氧)
- a heme copper protein 血红素铜蛋白 // an integral membrane protein 膜整合蛋白
- subunit that contains oxygen binding site has twelve membrane spanning domains 包含氧气结合位点和12个跨膜结构域
- a redox driven proton pump 氧化还原驱动质子泵
-
bd-type (稳定期、低氧)
- Not structurally related to the bo-type or mitochondrial type oxidases 与bo型在结构上不相关
- unique to bacteria 细菌特有的 // a very high affinity for O2 对氧气有很高亲和力
-
-
正向电子传递【电子传递链的标准形态】
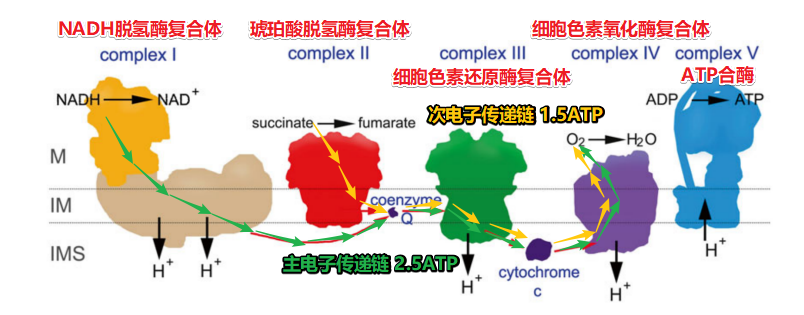
-
从NAD+/NADH,黄素单核苷酸FMN,辅酶Q,细胞色素bc1, 细胞色素c, 细胞色素aa3,还原电势越来越高
电子传递系统分类和举例
自然界存在多种多样的基于电子传递链的化学渗透系统,与生物的生存环境和适应机制紧密相关
超短电子传递链(不涉及脂溶性的膜上电子载体 )
-
重要的特点
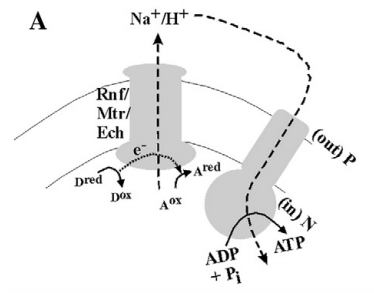
- 出现在大多数的产甲烷菌和产乙酸菌中,其利用H2还原CO2放出能量
- 标准的ΔEm仅为180 mV和130 mV,其产生的能量十分贫乏
- 仅仅包含一种化学渗透耦合的酶(在产乙酸菌中,该酶称为Rnf-复合物,在产甲烷菌中,该酶是一种甲基转移酶)
-
几个具体的案例
-
标准模式图
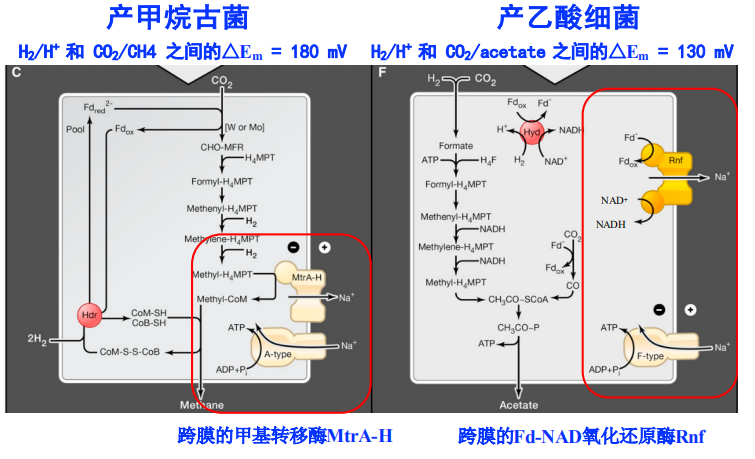
-
不含细胞色素的产甲烷古菌
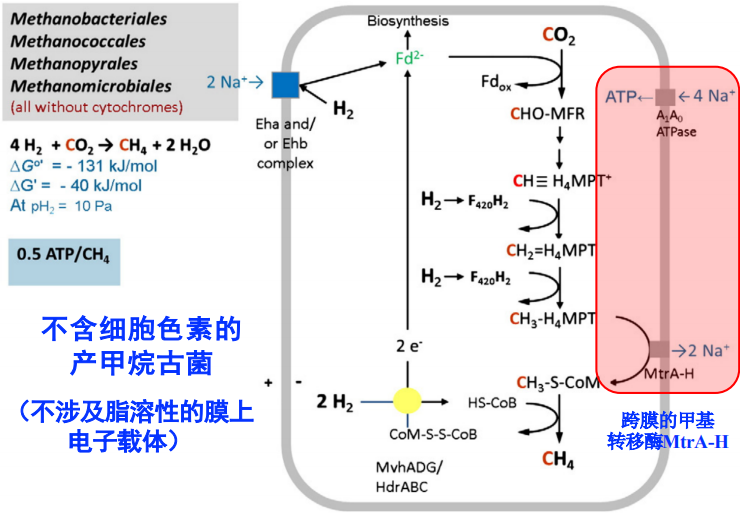
- 利用跨膜的甲基转移酶MtrA-H,每转移1个甲基泵出2个Na+
- 利用A1A0 ATPase合成ATP,每4个Na+合成1个ATP
- △E_m = 180 mV
- 不涉及脂溶性膜上的电子载体
-
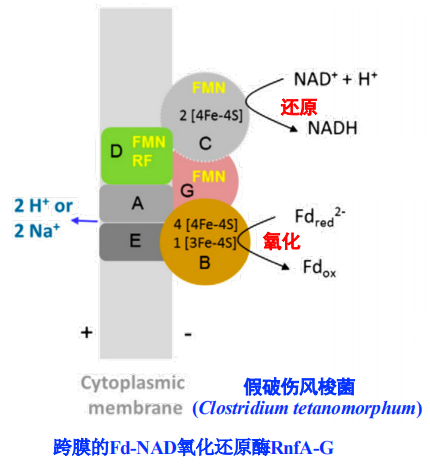
不含细胞色素的产乙酸细菌
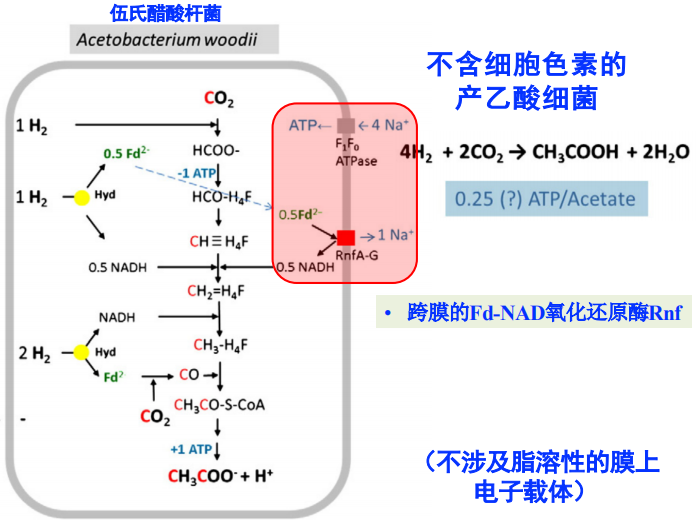
- 跨膜的Fd-NAD氧化还原酶Rnf
- 利用H2还原的Fd2-来驱动,每分子H2还原0.5Fd2-,泵出1个Na+
- △E_m = 130 mV
- 改过程亦步设计脂溶性膜上的电子载体
- 关键的氧化还原酶RnfA-G的结构
-
克氏梭菌的乙醇-乙酸发酵
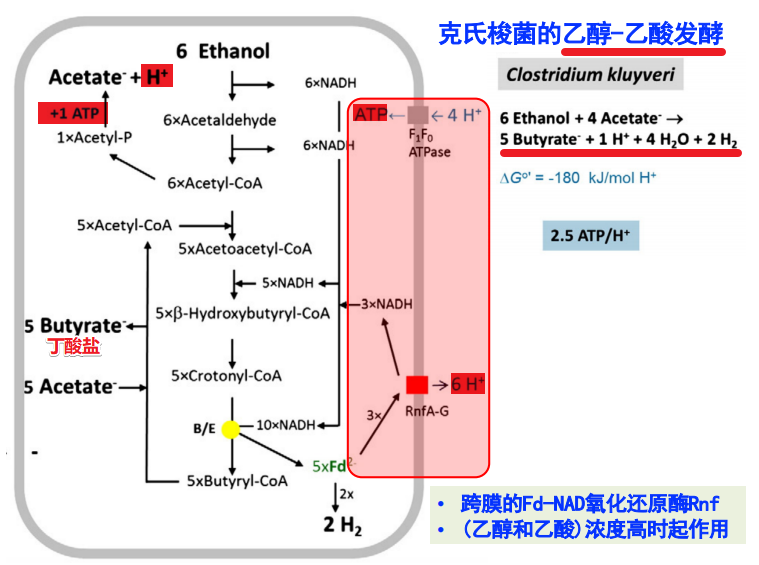
- 跨膜的Fd-NAD氧化还原酶Rnf,在乙醇和乙酸浓度高时起作用
- 其利用乙酸/乙醇发酵过程中产生的NADH来驱动RnfA-G,泵出H+
- 随后利用F1F0-ATPase产生ATP // 2.5ATP/H+
-
强烈火球菌的葡萄糖发酵(超嗜热古菌)
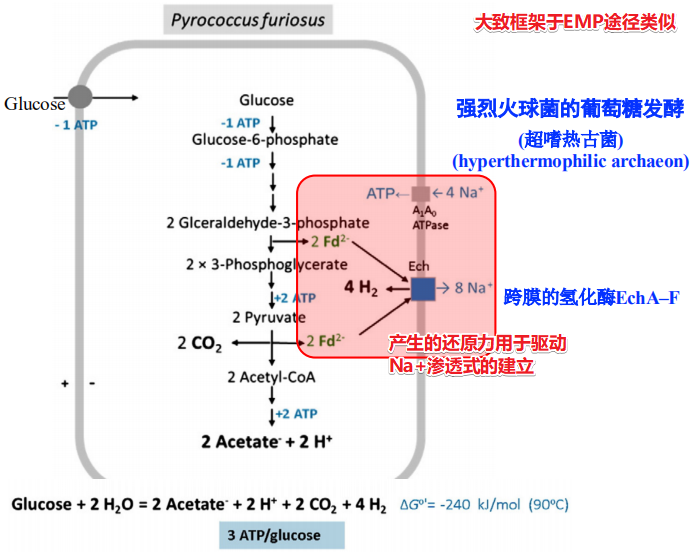
- 产生的还原力Fd2-用于驱动Na+的外流
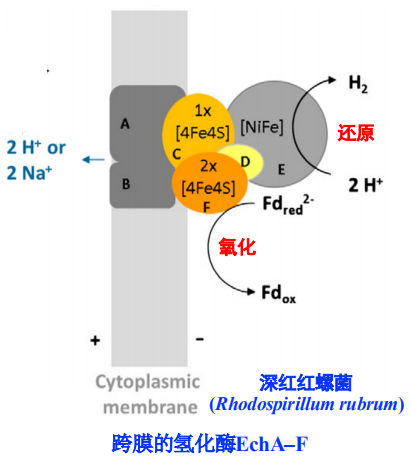
- 使用单个跨膜的氢化酶来建立化学渗透梯度
- 跨膜的氢化酶EchA-F的结构
-
涉及脂溶性的膜上电子传递
-
细胞捕获能量(节约消耗)的手段——图仅为关于原理的构象(可能的三种基本模式,不一定是真实的!)
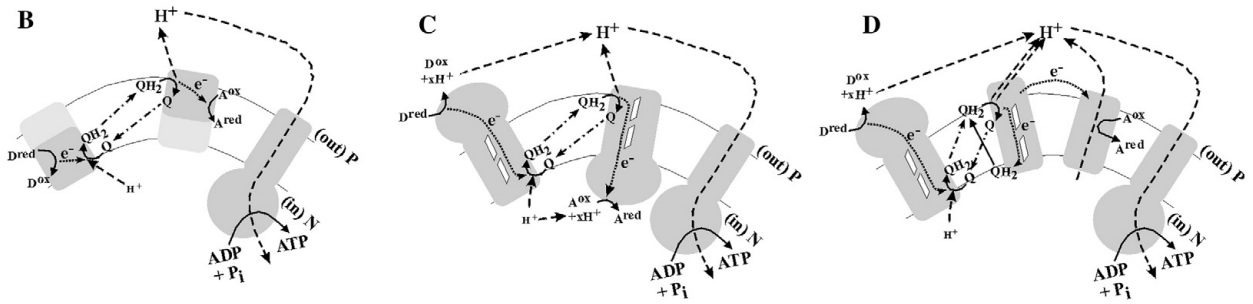
-
关键问题 :细胞如何产生更多的能量? 更优的底物;如何捕获并利用更多的能量? 建立更高的质子梯度,利用更充分的底物!- 使用氧化还原电势相差更大的底物,从根本上获得更多的化学能释放<从原始的H2/CO2到有氧呼吸,其底物电势相差越来越大>
- 利用阶梯还原的策略,使得物质中的能量可以被充分利用<例如反硝化作用,从NO3-到N2需要好几步>
- 利用氧化还原的复合体偶联质子泵(通过构象变化泵出更多质子) <例如:complex I的双稳态机制、Heme-copper O2 reductase>
- 利用电子传递体的不对称运送,泵送出更多的质子<例如:利用各种醌类物质和亚甲基吩嗪的反应位点和电子供体D不在同一侧,利用两个Heme在中间传递电子>
- 在供体氧化复合物与受体还原复合物中间加入其他酶 <例如:Rieske/cytb 复合物 涡轮增压 (包括 bc1-type (complex III) 和 b6f-type 复合物)>
-
B图:相比于超短电子传递链,若能将氧化和还原过程分开进行,中间利用合适电位的电子载体连接,即可以利用电子载体结合氢而不改变电位的性质,运送更多质子出膜,建立更高的质子梯度
-
C图:因为氧化过程往往伴随着质子的释放,而还原过程往往伴随着质子的结合,若可以对氧化和还原过程发生的位置加以控制,则可以在底物反应的过程中建立质子梯度
-
D图:Rieske/cytb complexes可以将QH2结合的质子释放,从而使其能够结合并运送更多的质子,建立更高的质子浓度梯度
-
-
脂溶性电子载体:醌类和亚甲基吩嗪(在膜上的不对称作用模式是重点!)
-
主要有两类物质,Quinones (醌类:包括mena-, ubi-, plasto-, 和caldariellaquinones)和Methanophenazine (亚甲基吩嗪)
-
这些载体共转移两个电子和两个质子
-
作用模式:可以在膜内扩散(diffusion):通常Q的还原位点位于细胞质一侧 (N),QH2氧化发生在另一侧(P)
- 电子供体D与Q的各自反应位点不在同一侧,两个血红素(heme)在中间传递电子
- 电子受体A与QH2的反应位点不在同一侧,两个血红素(heme)在中间传递电子
-
在细菌和古菌中的分布
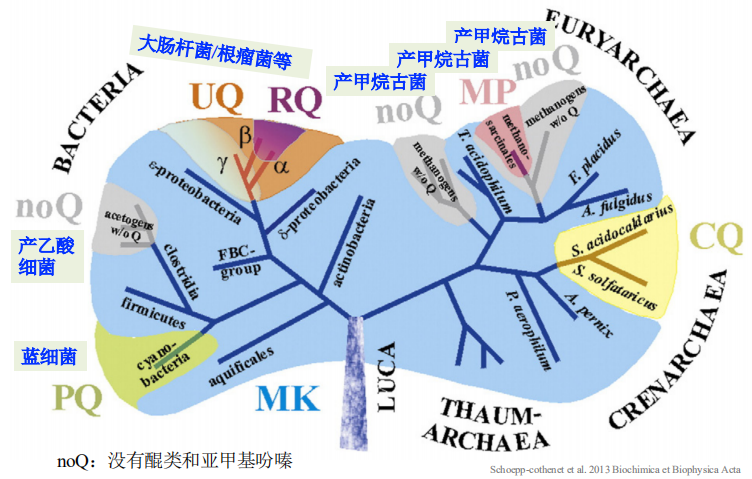
-
根据其电势的高低,可以初步将其分为两大类
- 高电势的:UQ、PQ、CQ
- 低电势的:MK、RQ
Complex I 的双稳态机制 基本概念(complex I是个啥,一些基本信息 http://www.complexi.org/overview.html ) Mitochondrial Complex I is the only enzyme that can take these electrons from NADH and derive energy stored in there. Complex I是细胞内唯一可以转化NADH为能量的酶! The mitochondrial enzyme is a huge molecule composed of at least 45 (!!) different proteins. 由至少45个蛋白组成的巨大复合体!(7种由线粒体基因编码) Coupled to the electron transfer, protons are pumped from the matrix side to the intermembrane space of mitochondria. 结合电子转移,质子从基质侧泵送到线粒体的膜间空间 Its’ L-shaped molecule sits in the membrane so that one arm is exposed outside and another arm is embedded into the membrane. L形状的分子,一端嵌入到膜中! This enzyme complex contains a noncovalently-bound FMN molecule and 2 binuclear and 6 tetranuclear iron-sulfur (FeS) clusters, of which 2 binuclear and 4 tetranuclear clusters are EPR-visible. 该酶复合物包含一个非共价结合的FMN分子和2个双核和6个四核铁硫(FeS)簇,其中2个双核和4个四核簇对EPR可见
双稳态机制!!! 基本模块组成 模块N:NADH氧化 FeS簇:Fe2S2 与 Fe4S4(FeS簇数量在部分原核生物有区别);连接NADH氧化位点与ubiquinone还原位点 模块Q:ubiquinone还原 模块P:质子泵 双稳态模型 E-state:(electron transfer) 电子传递 电子高速地从FeS簇N2传给泛醌或半醌,但是底物被遮蔽从而避免质子化; 泛醌的阴离子氧化还原中间体不稳定,质子泵处于input构象 P-state:(protonation) 质子化 电子不能从FeS簇传给泛醌或半醌,但是后者能被质子化; 泛醌的阴离子氧化还原中间体稳定,质子泵处于output构象Complex III的涡轮增压机制 发生于线粒体complex III上,利用反向电子传递实现了QH2的循环利用 QH2被氧化为Q而可以回收更多上游氧化还原反应所释放的能量
经典案例 甲烷八叠球菌的产甲烷途径(含脂溶性电子载体 亚甲基吩嗪) 代谢的全局图 脂溶性载体的局部图
反硝化作用(denitrification) // 硝酸盐呼吸 基本特点 又称为硝酸盐呼吸,以NO3-代替分子氧作为电子受体的无氧呼吸作用 完整还原过程的产物为氮气 --- 在低氧或无氧条件下进行 ATP产率降低,但是可以存活与繁殖 基本过程 硝酸还原酶的结构给个解释图(分成几个部分,核心的FeS簇在NarG上) 整合在膜上的 NarI 在膜的两侧传递电子(上面有两个Heme在膜的两侧) 大肠杆菌的硝酸盐呼吸 厌氧氨氧化 厌氧条件下,以亚硝酸为电子受体,以氨为电子供体产生氮气的反应 氨与一氧化氮在厌氧氨氧化细菌的联氨合酶及联氨脱氢酶催化下,氧化为氮气 基本过程
反向电子传递 电子歧化反应 为什么需要反向电子传递 在醋酸杆菌中,正常情况下可以利用H2还原CO2生成Fd2-从而建立Na+浓度梯度,进行产生ATP 但在依赖环境中低能底物(乳酸/乙醇)时,底物水平磷酸化可以产生ATP,但是却不能形成Fd2-,从而Na+梯度被破坏 反向电子传递,就是要依靠底物水平磷酸化产生的ATP和NADP来促进Fd2-的形成!
什么是反向电子传递 电子的传递能量爬坡,向氧化还原电势更负的电子传递链成分传递 如上例中,需要利用NADP(Em=-320mV)来促使Fd2-形成(Em=-420mV) 电子歧化反应(Electron bifurcation)和反向化学渗透是实现反向电子传递的两种手段 从生物能学角度看,电子歧化反应比ATP水解或反向化学渗透更经济,节约了ATP 什么是电子歧化反应 两个电子流向两个不同的单电子受体:一个电子受体具有较高的还原电势,另一个电子受体具有较低的还原电势(与供体氧化还原电对相比) 一个解释图,注意上面的电子载体的电势信息 电子歧化反应有两种类型 Flavin-based 电子歧化:四个不相关的蛋白家族,在厌氧细菌和厌氧古菌中存在,主要利用细胞质蛋白 Quinone-based电子歧化:三个酶结构与功能类似,在三域细胞生物中存在,主要利用膜蛋白 电子歧化酶的共同特征 歧化酶辅因子(Flavin和Quinone)都具有三个连续的氧化状态: ox = 0, ox = -1, ox = -2 Crossed-over redox potentials: the quinone/semiquinone (Q/SQ) couple has a more negative midpoint potential than the SQ/hydroquinone (SQ/HQ) couple 醌Q/半醌SQ比半醌SQ/氢醌HQ具有更负的中点电势 ∆E = Em(SQ/HQ) – Em(Q/SQ) >0 Quinone-based电子歧化(主要说Complex III上的Q cycle) 基本过程(电子的转移过程) 1./4. QH2释放 1st 电子给氧化状态的Rieske[2Fe2S](吸能),自身氧化为SQ; 2./5. Q/SQ电对具有 more negative 还原电势,能释放 2nd 电子给Cytochrome bL(放能); 以传递1st电子的能量消耗为代价,放大另一个电子的还原力 电势的变化图 Flavin-based电子歧化(有4类) electron-transferring flavoproteins (EtfAB/CarED/LctCB/ FixBA homologs) (EtfAB) -Bcd complex系统 (EtfAB/CarED) -CarC complex系统 膜固定的 (EtfAB/FixBA) -CX complex系统 案例:伍氏醋酸杆菌(利用NADH转化为Fd,用于构建Na+浓度梯度)(相当于一种将NADP转化为ATP的反应体系) NAD(P)H dehydrogenase (NuoF homologues) 电子歧化NADH脱氢酶-氢化酶 Electron-bifurcating dehydrogenase-hydrogenase 案例:伍氏醋酸杆菌 NADH-dependent ferredoxin:NADP reductase (NfnAB) heterodisulfide reductase (HdrABC) or HdrABC homologues 电子歧化异二硫化物还原酶-氢化酶 MvhADG-HdrABC Electron-bifurcating heterodisulfide reductase-hydrogenases 案例:Fd作为早期的能量通货? Just as ATP is an ideal energy currency that can couple exergonic and endergonic reactions, the same is true for Fdred, but this is restricted to redox reactions. 反向电子歧化(归中!) #QUESTION 在11个实验验证的Flavin-based电子歧化反应里,有5个是可逆的 反向电子歧化反应间接增加了厌氧微生物的细胞能量 案例:反向电子歧化反应有助于利用低能量底物(乳酸)生长 -
另外,之前在生化课上学习氧化呼吸链时,总是记不住,为了方便速查,借别人的图汇总过一张图还挺不错的。
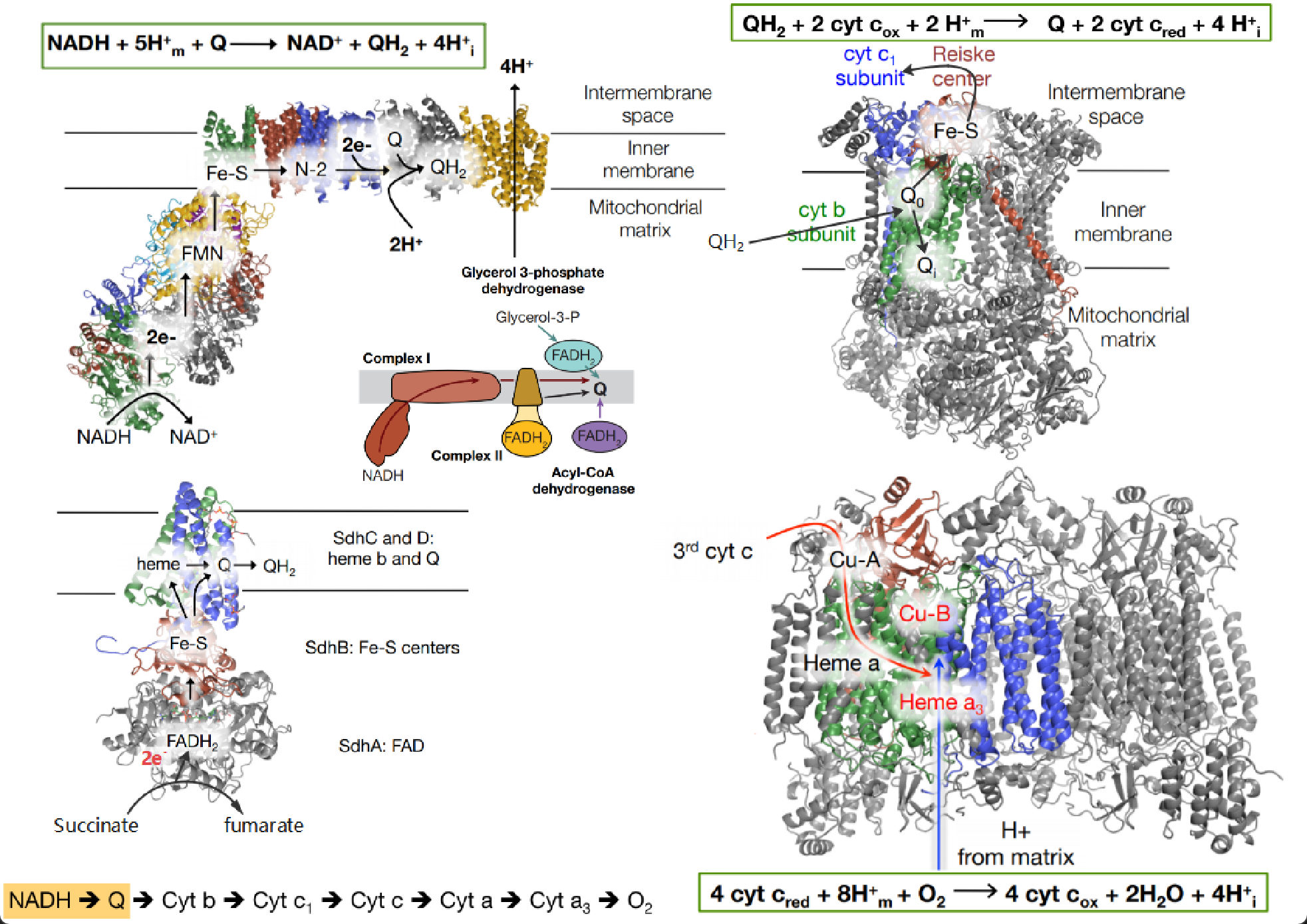 (2018年12月9日)
(2018年12月9日)
